Последствия внедрения: Недотрога желёзконосная
Impatiens glandulifera Royle
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Хотя в качестве декоративного растения I. glandulifera выращивали в садах Европы с середины XIX века, а в Северной Америке — с начала XX века, первые случаи дичания вида отмечены через 20 смен поколений, внедрение в естественные ценозы — через 60, а бурная экспансия — через 100 смен поколений.
I. glandulifera встречается на побережьях озёр и морей, на лугах, по рекам и ручьям, в зарослях чёрной ольхи и в лесах около селений [Kurtto, 1996]. Отмечено, что во влажных лесах вид занимает «новые» местообитания, то есть является пионерным. В Швеции заросли I. glandulifera сохранялись в течение по крайней мере 70 лет [Larsson, Martinsson, 1998]. При некоторых обстоятельствах вид только сокращает численность аборигенных видов, которые все же находят достаточно мест для произрастания, например в высоко динамичном пойменном лесу [Kasperek, 2004]. Но в иных ситуациях растение формирует плотные заросли, которые покрывают всю почву и «душат» другую растительность, причём I. glandulifera конкурирует не только с однолетними, но и с многолетними видами растений [Larsson, Martinsson, 1998]. В окрестностях Осло (Норвегия) отмечалось, что распространение I. glandulifera уменьшило видовое разнообразие в некоторых локальных ценозах. В речных долинах I. glandulifera формирует плотные заросли собственного сообщества, которое классифицируется фитосоциологами как ассоциация Impatienti-Calystegium [Moor,1958; Soo, 1971]. Способность I. glandulifera к конкуренции с природной флорой и её предсказуемая экспансия по водным путям указывают, что в будущем вид может стать более серьезной угрозой сохранению естественного биоразнообразия [Prach, 1994; Pysek, Prach, 1995]. Известно, что I. glandulifera предотвращает возобновление лесов в сырых полузатенённых местообитаниях [Lhotska, Kopecky, 1966].
Поскольку I. glandulifera, по-видимому, положительно реагирует на увеличение CO2 и повышение температуры, потенциально вид может стать в условиях изменяющегося климата ещё более агрессивным захватчиком.
I. glandulifera привлекает опылителей, и вид, по-видимому, способен конкурировать с другими видами Impatiens за опылителей [Daumann, 1967]. Путём конкуренции за опылителей I. glandulifera может заместить и некоторые другие природные виды [Chittka, Schurkens, 2001].
Генетические изменения и вариабельность вида. В естественном ареале в Кашмире и Джамму 2n = 20 [Khoshoo, 1957]. Во вторичном ареале вид существует по меньшей мере в двух формах, имеющих 2n = 18 и 2n = 20, причём первая считается производной, возникшей на новой родине [Khoshoo, 1957]. Популяция растений с 2n = 18 и n = 9 обнаружена в Манчестере [Valentine, 1971]. А в Кью обе формы произрастают вместе в одной и той же микропопуляции [Jones, Smith, 1966].
И в естественном, и во вторичном ареале отмечен полиморфизм по окраске цветков I. glandulifera. Основные цвета: лиловый, бледно-розовый и белый. Уже при первичной интродукции в Англию в 1838 году завезена не только лилово-, но и белоцветковая форма. Однако натурализовавшиеся растения с белыми цветами появились в Британии только в 1921 году [Valentine, 1971]. В континентальной Европе эта форма встречается только в культуре: отмечена в Цюрихе в 1914 году [Hegi, 1923] и на юге Польше [Valentine, 1971]. Выращивается форма и в канадской провинции Онтарио [Scoggan, 1978]. В СССР форма с белыми цветками также отмечена только в культуре — собрана А. К. Скворцовым в городе Мукачево (Закарпатье) и Д. Басаргиным [1989] на Дальнем Востоке.
Бледно-розовые цветки встречаются редко. В Канаде такая форма отмечена в провинциях Новый Брансуик, Британская Колумбия, Онтарио и Новая Шотландия [Scoggan, 1978]. На Дальнем Востоке такая форма встречается только в садах, а в европейской части России — и в спонтанных популяциях.
По данным D. Valentine [1971], только одна популяция натурализовавшейся I. glandulifera около города Манчестер и одна — на севере Голландии имеют больший процент светло-розовых и белых цветков, чем тёмно-пурпурных. Во всех остальных европейских популяциях наблюдается обратное соотношение.
Одной из причин узкого распространения белых цветков считалось то, что бледно-цветковые растения продуцируют меньше нектара и реже посещаются шмелями и пчёлами, чем тёмные цветы в соотношении 1:2 или 1:3 [Valentine, 1971]. Однако на экспериментальном участке ГБС РАН в интродукционной популяции, созданной из семян, собранных в различных точках вторичного ареала вида, все цветовые формы насекомые посещали одинаково часто, а бело-цветковые формы завязали такое же количество семян, что и лилово-окрашенные. При свободном переопылении различно-окрашенных форм в первой генерации произошло расщепление у потомства семян, собранных с бело-цветковых форм, в соотношении 2 пурпурных: 2 розовых: 1 бело-цветковое растение; в последующих генерациях число особей с белыми цветками ещё более сократилось [Виноградова, 2008].
Таким образом, в Англии и Западной Европе, где I. glandulifera натурализовалась ранее, наблюдается 5 вариаций окраски лепестков, причём голубые цветки возникли, по-видимому, уже во вторичном ареале. В районах, где I. glandulifera появилась недавно, форм с голубыми цветками не найдено вовсе, белые и светло-розовые имеются только в садах, а дичают экземпляры с тёмно-пурпурными и розовыми цветками.
Анализ дискретных признаков эстивации (взаимного расположения лепестков зигоморфного цветка) показал слабое внутрипопуляционное разнообразие [Басаргин, 1989].
Экономическое и социальное (положительное/отрицательное) значение. Уничтожение зарослей — дорогостоящий и долговременный процесс из-за высокой способности вида к восстановлению и распространению. Агентство по охране окружающей среды Великобритании подсчитало, что уничтожение I. glandulifera только в Англии и Уэльсе будет стоить от 210 до 240 миллионов евро.
Растение популярно среди коллекционеров бабочек, так как вид является важным источником нектара и пыльцы. Цветы часто посещают пчёлы и шмели [Beerling, Perrins, 1993]. Кроме того, I. glandulifera считается красивым декоративным растением; его семена продолжают высевать, и это играет важную роль в распространении вида.

Внедрение Impatiens glandulifera в прибрежные сообщества, Московская область, река Москва в Одинцовском районе
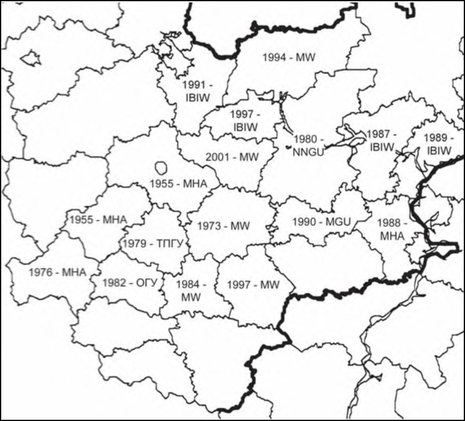
Расселение Impatiens glandulifera в Средней России

Массовое цветение Impatiens glandulifera в Москве, в овраге в Битцевском лесопарке

Розовая окраска венчика Impatiens glandulifera

Дурнишник эльбский
Xanthium albinum (Widder) H.Scholz

Недотрога мелкоцветковая
Impatiens parviflora DC.




