Таксономические проблемы изучения инвазионных видов
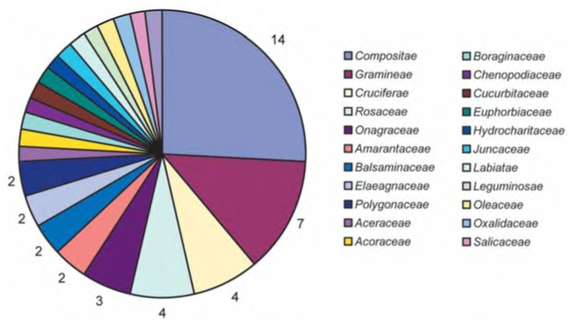
Таксономическое распределение инвазионных видов флоры Средней России таково: несомненный лидер — семейство Compositae с 14 видами, на втором месте Gramineae — 7 видов, далее следуют семейства Cruciferae и Rosaceae — по 4 вида и Onagraceae — 3. Остальные семейства, среди которых Асеrасеае, Асогасеае, Amaranthaceae, Balsaminaceae, Boraginaceae, Chenopodiaceae, Cucurbitaceae, Elaeagnaceae, Euphorbiaceae, Hydrocharitaceae, Juncaceae, Labiatae, Leguminosae, Oleaceae, Oxalidaceae, Polygonaceae, Salicaceae и Umbelliferae, представлены 1-2 видами.
Таксономическое распределение инвазионных видов флоры Средней России
Отметим, что эта таксономическая структура соответствует пропорциям адвентивной флоры Европы, выявленной в результате паневропейского проекта DAISIE [Lambdon et al., 2008]. Различия невелики: 3-е место в адвентивной флоре Европы занимает Rosaceae, заметно опережая семейство Cruciferae. Это отличие легко объяснимо. Среди среднерусских адвентивных растений немало широко распространённых видов розоцветных (Malus domestica, Primus domestica, Pyrus communis и другие), которые не являются инвазионными видами. Крестоцветные, напротив, отличаются высокой активностью и широко расселяются из мест заноса.
Интересно, что таксономический спектр адвентивного и инвазионного компонентов флоры Средней России имеют незначительные отличия, главным образом, количественные. В адвентивном компоненте первые пять семейств располагаются следующим образом: Compositae (13%) — Gramineae (13%) — Cruciferae (9%) — Rosaceae (8%) — Leguminosae (7%). Инвазионный компонент флоры Средней России даёт следующий таксономический спектр: Compositae (27%) — Gramineae (10%) — Cruciferae (8%) — Rosaceae (6%) — Onagraceae (6%). [Виноградова, Майоров, Хорун, 2006].
Гораздо более существенные различия отмечены для таксономических спектров кенофитов в других регионах. Так, спектр кенофитов Украины: Compositae (32%) — Gramineae (8%) — Cruciferae (8%) — Umbelliferae (6%) — Amaranthaceae (6%), а спектр кенофитов Польши Compositae (24%) — Cruciferae (9%) — Leguminosae (8%) — Gramineae (5%) — Scrophulariaceae (5%).
Впрочем, не стоит переоценивать важность таксономического распределения для анализа парциальных флор при небольших выборках. При разнообразных подсчётах следует учитывать статистический характер выделенных групп и неизбежные разночтения или даже ошибки [Щербаков, 2001, 2003, 2006]. Более того, на таксономический спектр адвентивной флоры влияет степень ее выявления; в отличие от природной флоры, эти пропорции по мере накопления данных изменяются [Щербаков, 2008]. Следует иметь в виду, что традиционно в российских флористических исследованиях используют систему А. Энглера (с изменениями!), западные флористы же постепенно переходят на систему APG (the Angiosperm Phylogeny Group), в том числе и при исследовании адвентивной флоры [сравни: Маевский, 2007 и Lambdon et al., 2008].
Особую важность при работе с адвентивными растениями имеет надёжная идентификация видов, так как вся необходимая информация (происхождение, тип ареала, экологические свойства и тому подобное) «привязаны» к названию таксона. Без точного определения инвазионных видов объединение усилий исследователей неэффективно [Pysek, 2003; Pysek et al., 2008]. Это определение необходимо для принятия ряда управленческих и экономических решений по контролю над инвазионными видами [Smith et al., 2008].
Точное определение заносных видов наталкивается на ряд объективных проблем:
- Новые для данной территории адвентивные виды часто отсутствуют в доступных региональных «Флорах» и определителях;
- Гербарные образцы из «иностранных» отделов Гербариев (MW, МНА и даже LE) обычно немногочисленны и часто неоднородны из-за заметного числа неправильных определений
- Адвентивные виды нередко представлены нетипичными, уклоняющимися формами;
- Адвентивные виды, особенно эргазиофиты, часто несут черты предка-основателя.
Все это существенно затрудняет надёжную и точную идентификацию. Поэтому в истории изучения адвентивной флоры немало случаев номенклатурной и таксономической путаницы. Приведём несколько примеров.
Род бруннера (Brunnera Steven) на территории России представлен двумя видами: кавказской бруннерой крупнолистной В. macrophylla I. M. Johnst. и бруннерой сибирской В. sibirica Steven [Попов, 1953]. Как декоративное растение в наших условиях выращивают преимущественно В. sibirica с более крупными цветками и лучшими декоративными свойствами [Чуб, 2008]. Это неприхотливое растение подолгу удерживается на местах прежней культуры и встречается как заносное у дачных посёлков, по сорным местам. Однако во «Флоре европейской части СССР» для нашей территории указывается лишь B. macrophylla [Доброчаёва, 1981]. Вслед за этим бруннера крупнолистная появилась в ряде региональных работ [например, Цвелёв, 2000; Григорьевская и другие, 2004]. Таксономическое решение Д. Н. Доброчаевой основано, по-видимому, на «Flora Europaea» [Chater, 1972]. Рисунок в известном определителе Стейси подтверждает, что, по крайней мере, в Великобритании культивируется именно B. macrophylla [Stace, 1997]. Среди адвентивных растений Европы значится только последний вид [Handbook..., 2009]. Однако описание бруннеры, данное Д. Н. Доброчаевой [1981], соответствует B. sibirica у М. Г. Попова [1953].
Наши наблюдения за природными популяциями B. macrophylla на Кавказе, а также за культивируемыми растениями в Ботаническом саду МГУ и подробное изучение гербарных материалов с Кавказа и из Сибири (MW, МНА) показали, что как декоративное растение культивируется именно бруннера сибирская. B. macrophylla отличается от B. sibirica относительно мелкими цветками, наличием эпигеогенных корневищ и более ранним цветением: цветки у неё раскрываются ещё до полного разворачивания крупных прикорневых листьев. Кроме того, у B. macrophylla корневище сравнительно тонкое [Чуб, 2008]. Следует помнить, что размер цветков не является надёжным диагностическим признаком [Попов, 1953]. B. macrophylla отмечена в Подмосковье лишь в последние годы (личное сообщение В. Д. Бочкина).
Таким образом, таксономическая обработка Brunnera в последней по времени крупной региональной сводке не описывает реальной ситуации с видами рода на нашей территории. Поэтому без обращения к гербарным материалам литературные указания следует воспринимать как предварительные данные, поскольку не ясно точно, о каком именно виде идёт речь.
Для викарных видов в пределах природного ареала нередко самым надёжным «признаком» является географическое положение популяции. Так, европейско-кавказский Acer tataricum L., среднеазиатский A. semenovii Regel et Herd, и дальневосточный A. ginnala Maxim, аллопатричны и достаточно хорошо отличаются морфологически [Цвелёв, 2000]. Однако, в последнее время многие европейские ботаники [см., например, Suomen..., 1992; Zander, 2002] объединяют эти виды. Причиной такого мнения может быть как широкое распространение в Европе сортовых форм с промежуточными признаками, так и недостаточно точное морфологическое описание во «Флоре СССР» из-за невнимания к гетерофилии у этих видов на генеративных и вегетативных побегах [Пояркова, 1949]. A. tataricum и A. ginnala широко используются в озеленении, в последнее время неоднократно отмечается самосев. Однако, как оценивать необычные формы клёнов из культуры с промежуточными признаками не ясно: либо это результат сортового отбора уклоняющихся форм одного из видов, либо гибридизация близких видов при вторичной симпатрии. Современные методы в ряде случаев способны определить географическое происхождение инвазионных растений [смотрите, например: Milne, Abbott, 2000; Meekins et al., 2001; Durka et al., 2005; Kliber, Eckert, 2005].
Особую проблему представляют гибридные комплексы. Так, в культуре широко распространены близкородственные виды — дальневосточная Spiraea salicifolia L. с розовыми цветками и компактным цилиндрическим соцветием и североамериканская S. alba L. с белыми цветками и пирамидальным соцветием. Время от времени сообщается о находках этих спирей как заносных растений. Кроме того, в культуре несколько реже встречается североамериканская S. douglasii Hook, с интенсивно-розовыми цветками, компактным соцветием и обильно опушёнными снизу листьями. Известны, описаны и широко культивируются гибриды между этими тремя видами. Следует учитывать, что иногда крупные формы S. alba выделяют как S. latifolia Aiton, а в составе S. douglasii известны и описаны гололистные растения — это ssp. menziezii (Hook.) Voss. Некоторые гибриды «S. latifolia» описаны как особые таксоны [Цвелёв, 2000]. Очевидно, что в подобных случаях надёжная идентификация гибридных образцов едва ли возможна, поскольку ситуация осложняется селекцией высокодекоративных форм спирей, отбор направлен на окраску цветков и форму соцветий — диагностические признаки у родительских видов и гибридных форм.
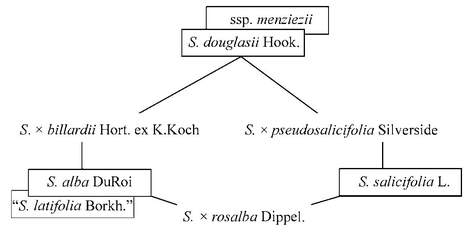
Спиреи S. salicifolia, S. alba, S. douglasii и их гибриды
Определение гибридных растений наталкивается на ещё одну диагностическую проблему. Нередко гибридизация происходит между растениями в условиях культуры вне природного ареала. Это, например, Reynoutria х bohemica, S. х uplandicum Nyman (S. officinale x S. asperum Lepechin), Aster x salignus и, вероятно, Amelanchier spicata. В этом случае такие гибридные формы часто неизвестны из мест симпатрии родительских видов (если эти ареалы пересекаются) и, естественно, отсутствуют в региональных «Флорах...» и определителях.
Не всегда удаётся установить оба родительских вида для гибридного таксона. Так, ирония Мичурина, или черноплодная рябина, — Aronia х mitschurinii A. K. Skvortsov et Yu. K. Maitulina была получена при селекции A. melanocarpa (Michx.) Nutt. et Elliott. Для этого И. В. Мичурин использовал гибридизацию с другими видами, в том числе, вероятно, рябинами. К сожалению, история возникновения «черноплодки», несмотря на обширные публикации работ Мичурина, не ясна. Следует учитывать, что для гибридизации он нередко применял смесь пыльцы разных растений, что, по мнению селекционера, должно было повышать качество гибридного потомства. А. х mitschurinii отличается от A. melanocarpa, с которой её нередко путают, прежде всего большей мощностью растения и качеством плодов [Скворцов, Майтулина, 1982]. Точное установление второго родительского вида возможно только при применении методов молекулярной генетики.
В 1970-1980-е годы аронию широко культивировали в промышленных масштабах как неприхотливое высокоурожайное ягодное растение, плоды которого, среди прочего, выделяются высоким содержанием витамина Р. В последние годы отмечено расселение аронии Мичурина по лесным опушкам и на вырубках, чаще у дачных посёлков или близ плантаций черноплодки. При этом она нередко поселяется по сырым местам вдоль дорог, на вырубках, то есть в местообитаниях аналогичных условиям произрастания A. melanocarpa в естественном ареале.
В некоторых случаях адвентивные растения показывают иной характер изменчивости, чем в природном ареале. Так, кипреи из родства E. ciliatum Raf. представлены в Европейской России двумя видами: Е. adenocaulon Hausskn. и Е. pseudorubescens A. Skvortsov. На территории Северной Америки E. ciliatum разделяют на большое число микровидов, не всегда хорошо различимых. В Евразии, напротив, эти два кипрея ведут себя как независимые и вполне отчётливо различимые виды. Первоначально Е. pseudorubescens был определён как Е. rubescens Rydb., однако знакомство А. К. Скворцова с типовым материалом убедило его в ошибочности такого решения. Обилие числа описанных видов и малая доступность типового материала для кипреев не позволяет надёжно сопоставить адвентивный бело-цветковый кипрей и какой-либо североамериканский микровид. Это побудило А. К. Скворцова к описанию нового вида — Е. pseudorubescens. Не исключено, что в дальнейшем удастся обнаружить исходный североамериканский микровид из состава Е. ciliatum [Скворцов, 1996].
В средней полосе Европейской части России среди обычного тростника Phragmites australis (Cav.) Trin. ex Steud. как заносные отмечены очень крупные растения с широкими листьями, мощными соцветиями, зацветающие поздно осенью и размножающиеся, по-видимому, только вегетативно [Папченков, 2008]. Эти растения получили название Ph. altissimus Benth. (Ph. isiaca Delile). В настоящее время в средней полосе они известны в Ярославской, Тверской, Московской и Ульяновской областях, Мордовии [Папченков, 2008; MW]. Следует иметь ввиду, что эта раса хорошо отличается от типичной лишь во вторичном ареале. В обширном природном ареале наблюдается постепенный переход от растений тростника привычного нам облика к огромным южным формам. Н. Н. Цвелёв эту форму выделяет в ранге подвида ssp. altissimus (Benth.) Clayton, в целом более южного, но симпатричного с типовым подвидом [Цвелёв, 1974, 1976].
В ряде случаев идентификации трудно разграничимых видов помогает их биологическое окружение. Так, в Северной Америке галловая мушка Eurosta solidaginis предпочитает инфицировать Solidago altissima L., при этом близкий и очень похожий S. canadensis ею практически не повреждается [Abrahamson et al., 1989; Abrahamson; Weis, 1997]. Как это ни удивительно, наличие или отсутствие галлов — один из лучших диагностических признаков для определения этих близких видов золотарника [Semple, Cook, 2006]. Во вторичном ареале мы естественным образом лишены этого удобного признака.
Нередко в случае с адвентивными видами имеет место таксономические разночтения в трактовке тех или иных видов в зависимости от национальной школы. Например, в англоязычных странах дурнишник, называемый в нашей стране X. albinum, принято ошибочно объединять с X. strumarium. Или, американцы обычно называют адвентивный окопник из Европы Symphytum officinale L., между тем, большая часть образцов принадлежит к S. x uplandicum — US, NY, GH, А (определения С. P. Майорова и Д. Д. Соколова). Несомненны подобные случаи и в практике российских флористов. Примеры тому Cenchrus echinatus L. и C. pauciflorus Benth., Phytolacca acinosa Roxb. и Ph. americana L.
Многие используемые в озеленении древесные растения поступают из специализированных питомников или ботанических садов, где нередко близкие виды культивируются совместно. При этом в ряде случаев спонтанно возникают межвидовые гибриды. Такая гибридизация характерна, например, для многих розоцветных. Некоторые гибридные формы затем находят широкое применение в озеленении, некоторые из таких таксонов способны дичать. В нашем списке инвазионных видов к гибридогенным таксонам относятся Reynoutria х bohemica и Aster x salignus. По мнению некоторых учёных, такая гибридизация может приводить к образованию инвазионных нотовидов [Abbott, 1992; Ellstrand, Schierenbeck, 2000]. Поток генов между местными и инвазионными видами неоднократно доказан на генетическом уровне и требует тщательного изучения [Abbott et al., 2003; Petit, 2004; Bleeker, Matthies, 2005].
С другой стороны, гибридизация местных растений с адвентивными приводит к генетическому загрязнению, а интрогрессивная гибридизация может приводить к исчезновению редких местных видов [Bleeker, Matthies, 2005; Bleeker et al., 2007].
Следует отметить, что переход интродуцированного или заносного вида через географические и биологические барьеры [Richardson et al., 2000] видоизменяет инвазионный вид. Это было убедительно показано на гигантских борщевиках с использованием молекулярно-генетических методов: инвазионные популяции Heracleum mantegazzianum и Н. sosnowskyi обнаружили отчётливые отличия от природных [Jahodov6 et al., 2007; Ecology..., 2007]. Этот переход требует некоторого времени, в течение которого интродуцированный вид становится способным к активному расселению. Это время в англоязычной литературе принято называть «lag-phase». Для объяснения этого явления предложено несколько гипотез [обзор см.: Pysek, Hulme, 2005]. Нам ближе гипотеза о накоплении за это время генетических изменений, которые позволяют растению приспособиться к новым условиям и реализовать адаптивные преимущества адвентивного вида [Hobbs, Humphries, 1995; Kowarik, 1995].
* * *
Точная таксономическая идентификация адвентивных видов растений должна быть неотъемлемой частью любых работ, исследовательских или практических, с инвазионными видами [Smith et al., 2008]. Повысить точность определения можно лишь аккуратным изучением таксономической литературы, тщательным сравнением адвентивных растений с надёжно определёнными гербарными образцами из мест естественного произрастания. Таксономические комментарии должны сопровождать публикации, посвящённые адвентивным видам.
Остаются существенные проблемы в таксономии и диагностике адвентивных видов в родах Asters. 1. (Symphyotrichum), Helianthus, Crataegus, Rosa и тому подобного.
В нашем списке к таксонам с окончательно не решёнными таксономическими проблемами относятся Amelanchier spicata, Crataegus monogyna, Epilobium adenocaulon, E. pseudorubescens, Solidago canadensis, Aster x salignus, Helianthus tuberosum, Xanthium albinum. В очерках приводятся необходимые комментарии по систематике этих видов.
С другой стороны, Cardaria draba в настоящее время чаще включают в состав рода Lepidium, a Aster х salignus переносят в род Symphyotrichum. Мы сохранили привычные названия для этих растений, поскольку собственно границы видов не изменяются.
← Предыдущая глава