Вторичный ареал: Недотрога желёзконосная
Impatiens glandulifera Royle
Вторичный ареал
История интродукции и географическое распространение. Вторичный ареал вида очень широк и охватывает Северную Америку и Евразию. В Европе (Англия) I. glandulifera впервые появилась в 1838 году. Семена с 3 растений этого вида с лиловыми цветами и с 1 растения var. Candida с белыми цветами прислали в то время в Кью и начали разводить как декоративное растение. Позднее, в 1898 году, в Англию были завезены растения var. pallidiflora, которые встречаются в Сиккимских Гималаях [Valentine, 1971]. Упоминания о дичании растения в Британии долгое время были крайне редкими: один случай зафиксирован в 1855 году в Девоншире [Valentine, 1971], другой — в 1859 году около Манчестера, затем находки участились — в 1864 году I. glandulifera одичала в Ланкашире, в 1895 году в графстве Суррей, в 1898 году — в Вустершире [Britten, 1900].
В Германии I. glandulifera появилась, по одним данным, одновременно с появлением её в Англии [Valentine, 1971], а по другим — в 1897 году в окрестностях Гамбурга [Hegi, 1923]. Около 100 лет этот однолетник изредка культивировали в парках, на кладбищах, в сельской местности, порой возделывали как медонос. На юго-западе Германии в некоторых местах недотрога была довольно обычна уже в 1920-х годах, распространившись из швейцарских популяций (где вид натурализовался в 1904 году) по Рейну. Однако к 1923 году недотрога встречалась не более чем в 50 пунктах Средней Европы. Отмечалось, что I. glandulifera плохо закрепляется на одном месте, поскольку семена её полностью не вызревают [Hegi, 1923].
Вид был интродуцирован Ботаническим садом Хельсинки в конце 1800-х годов [Kurtto, 1992]. Первые натурализовавшиеся популяции наблюдались в Швеции в конце 1920-х годов, в Норвегии — в конце 1930-х годов, а в Финляндии — в 1947 году [Kurtto, 1996]. В Дании вид отмечен впервые в 1888 году, в 1956 году зарегистрировано уже — 40 находок, главным образом, в восточной части страны [Pedersen, 1956], а в 1988 году I. glandulifera произрастала уже по всей Дании [Hansen, 1991]. В Польше вид интродуцирован в Судетские горы в 1890 году. В Латвии I. glandulifera интродуцирована как садовое растение, а первый гербарный сбор датирован 1898 годом (RIG).
Расширение ареала I. glandulifera началось, по-видимому, после Второй мировой войны. В 1957 году недотрога найдена на берегу водоёма в Амьене, Франция [Wintrebert, 1957]. В 1968 году она отмечена в Хорватии, до 1989 года её обнаружили только 15 раз, а экспансия вида началась лишь в последнее десятилетие прошлого века [Lukac, 1989]. В 1979 году I. glandulifera отмечена близ города Павия в качестве нового для флоры Ломбардии вида [Soldano, 1979]. В начале 1960-х годов началась бурная экспансия вида в Средней Европе, и по сведениям E. Daumann [1967], в некоторых местах на юге Богемии I. glandidifera вытеснила аборигенную I. noli-tangere. В Литве вид как беглец из культуры зарегистрирован в 1959 году [Gudzinskas, 1998]. Первые популяции в естественных пойменных лесах по рекам Польши отмечены в начале 1960-х rодов [Jasnowski,1961; Dajdok, Aniol-Kwiatkowska, 1998].
Теперь I. glandulifera широко распространена в 18 странах Европы между 30° и 64° северной широты: в Ирландии, Англии, на юге и на восточном побережье Швеции, юге Норвегии и Финляндии, в Дании и Германии. Продолжается инвазия вида в Чехии и Словакии [Pysek, Prach, 1995], а также в Швейцарии, где вид широко распространился уже в поясе до 800 метров над уровнем моря. Как обычное растение недотрога желёзконосная отмечена во Французских Пиренеях, в Голландии, Австрии (особенно в Альпах), Польше, Венгрии, бывшей Югославии [Beerling, Perrins, 1993].
Спонтанные популяции обнаружены нами в 2003 году в Австрийских Альпах на берегу горной речки Мелль на высоте 900 м над уровнем моря, а на юге Австрии вид рос в придорожных канавах на протяжении 3-5 км, достигая в высоту 3 м. Во время экспедиционной поездки в Польские Татры (2005 год) мы наблюдали заросли недотроги желёзконосной, тянущиеся на километры. Обилие недотроги в странах Европы с каждым годом увеличивается, несмотря на проводимые гидромелиоративные мероприятия и применение гербицидов [Mountford, 1994].
В Северную Америку (Канада) недотрога попала, по-видимому, в 1906 году, изредка её культивировали, а в 1960-е годы в ряде штатов США (Калифорния, Айдахо, Массачусетс, Мэн, Мичиган, Монтана, Нью-Йорк, Орегон, Вермонт, Вашингтон) и в ряде провинций Канады (Британская Колумбия, Онтарио, Квебек, Нью-Брансуик и Новая Шотландия) недотрога начала внедряться в естественную растительность по берегам рек и по влажным местообитаниям [Gleason,1963; Scoggan, 1978].
Как интродуцированное растение отмечена во флоре Ирана [Grey-Wilson, 1979].
На территорию России I. glandulifera попала в конце XIX века. Известно, что в Петербургском ботаническом саду недотрогу культивировали в 1895 году, поскольку вид упомянут в списке семян сада [Delectus seminum..., 1895]. Сборы I. glandulifera в Курляндии сделаны в 1898 году (RIG). Случаи дичания вида на территории бывшего СССР были в то время, так же как в Средней Европе, крайне редки. К 1941 году известны лишь 2 местонахождения: окрестности озера Селигер [Маевский, 1940] и околицы города Каменец-Подольский (Украина), где недотрога найдена в 1939 году [Берко, 1963]. Во Флоре СССР указано [Победимова, 1949] ещё одно место сбора — Эстония. Однако и там вид встречался дико только в двух местообитаниях вблизи города Тарту рядом с ботаническими садами, где недотрога в изобилии культивировалась [Talts, 1959].
После войны зарегистрировано ещё несколько местообитаний недотроги: в Литве в 1959 году в одичалом состоянии [Gudzinskas, 1995] и на Украине: в 1953 году — в Прикарпатье, город Трускавец, в цветнике (LE); в районе города Ковель Волынской области [Карнаух, 1955] и в 1962 году в селе Верхняя Билка Львовской области в запущенном парке [Берко, 1963]. В 1956 году растение обнаружено в одичалом виде в Ленинграде [Гусев, 1973].
В 1960-1970-х годах бурная натурализация недотроги началась и в европейской части СССР: в 1967 году она отмечена в городе Приморск Ленинградской области, а ранее указана и для Петродворца [Гусев, 1973]. В 1974 году крупная популяция одичавшей недотроги отмечена в городе Рига (LATV). В 1978 году недотрога найдена Н. Н. Цвелёвым на левом берегу реки Уда на юго-западной окраине Харькова (LE). Массовое дичание I. glandulifera наблюдалось и в Удмуртии: А. Н. Пузырёв [1985] отмечает, что на небольшой речке у станции Можга цветущие растения образовали заросли по берегам на протяжении нескольких метров. В 1986 году найдена в Псковской области.
Ещё во второй половине 1920-х годов недотрога желёзконосная отмечена в городе Орджоникидзе и окрестностях [Горбачёв, 1932]. Но здесь она не распространялась, а возможно, и исчезла вовсе. Была приведена для окрестностей Кисловодска как первое указание для Кавказа в 1979 году [Котов, 1979]. В 80-х годах прошлого века найдена в регионе ещё в нескольких пунктах на рудеральных местообитаниях: единично на окраине посёлка Мизур в Алагирском ущелье в 1983 году, в окрестностях города Орджоникидзе в 1984 году, по склонам горы Лысая в Лесистом хребте в 1987 году [Комжа, Попов, 1990].
Довольно редки случаи разведения недотроги в Сибири. В 1988 году она отмечена В. В. Штаркером на приусадебных участках в районе заповедника Столбы. Через год найдена по берегам рек Танзыбей и Оя в Ермаковском районе Красноярского края [Степанов, 1990]. В 90-х годы прошлого века I. glandulifera стала дичать из культуры в Томской области [Пяк, 1994].
Во Владивостокском районе недотрога собрана В. Н. Ворошиловым как сорное растение в 1960 году (МНА). В 1974 году найдена Т. Н. Ульяновой в Южно-Сахалинске у забора (МНА) и в Корсаковском районе в сорных местах близ ж/д насыпей [Воробьёв и другие, 1974]. К 1980 году недотрога уже широко распространилась по огородам, садам и паркам города Хабаровск [Шлотгауэр, Небайкин, 1984], а в начале 90-х годов прошлого века натурализовалась на острове Кунашир [Нечаева, Петрова, 1995].
В настоящее время недотрога широко известна как агрессивный чужеродный вид в умеренных областях в Европе, Азии, Северной Америке и Новой Зеландии.
В более или менее естественных местообитаниях отмечается повсюду в Балтийском регионе [Kuusk et al., 1996]. В Финляндии с 1990-х годов расширение ареала I. glandulifera ускорилось, и теперь этот вид весьма обычен в плотно населённых районах на юге и в центре страны, а наиболее северные популяции достигли Северного полярного круга.
В Швеции недотрогу считают одним из пяти наиболее агрессивных видов растений. Она найдена по всей Швеции, кроме Лапландии [Larsson, Martinsson, 1998]. В Норвегии вид быстро распространяется, занимая естественные и полуестественные местообитания — сырые леса, сырые луга, затопляемые земли, канавы, побережья, речные потоки и различные типы сырых рудеральных участков; найден на юго-востоке Норвегии и по северу побережья до 69,5° северной широты [Aim, 2002; Lid, Lid, 2005]. В обоих ботанических садах Исландии вид отсутствует, и неизвестно, натурализовался ли он где-либо в Исландии [Nobanis, 2009].
В Литве I. glandulifera наиболее интенсивно распространяется по речным берегам, однако произрастает и во многих других типах естественных и антропогенных местообитаниях [Gudzinskas, Sinkeviciene, 1995; Gudzinskas, 1998], и её считают инвазионным видом.
Информации о расселении I. glandulifera в Латвии мало, и существует неоднозначное мнение о её агрессивности. Этот вид вторгся по берегам рек, найден в старых парках, садах, обочинах дорог и мусорных местах. Реки — главный путь распространения недотроги в Латвии [Nobanis, 2009].
В настоящее время I. glandulifera распространилась по всей Польше, но чаще встречается на юге, это один из 20 самых агрессивных инвазионных видов в стране [Tokarska-Guzik, 2003, 2005]. В Германии растение является наиболее широко распространённым чужеземным видом и растёт по берегам рек и другим влажным, богатым питательными веществами естественным местообитаниям. В Германии вопрос относительно агрессивности I. glandulifera дискуссионный, потому что нет доказанных случаев, когда опасности был бы подвергнут аборигенный вид. Расселение I. glandulifera ведёт к сокращению природных видов, однако в высоко-динамичных местообитаниях пойменных лесов всегда есть новые постоянно возникающие местообитания, и популяции I. glandulifera через какое-то время исчезают (например, в годы с неподходящими погодными условиями или при затоплении). Кроме того, вид преимущественно замещает Urtica dioica, которая не является редким или находящимся под угрозой исчезновения видом [Kasperek, 2004].
Пути распространения. Основной способ интродукции — «бегство» из частных садов и пасек. В Финляндии вид распространился, главным образом, при ввозе семян из-за границы и частном обмене семенным материалом. Часто распространяется также вокруг населенных мест из заброшенных садов или с почвой [Kurtto, 1996].
Статус в регионе. В одичалом состоянии недотрога впервые встречена близ Сенежского озера в Клинском уезде Московской губернии в большом количестве [Сырейщиков, 1914]. К 1941 году известно лишь ещё одно её местонахождение в окрестностях озера Селигер [Маевский, 1940]. В Москве и Калуге одичавшие экземпляры недотроги собраны в 1955 году (МНА). По личным сообщениям садоводов-любителей известно, что в конце 1950-х годов семена I. glandulifera, как декоративного растения, пользовались в Москве большим спросом.
В 1970-х годах началась бурная натурализация недотроги во всех областях Средней России. В 1971 году I. glandulifera собрана Ю. Д. Гусевым в Московской области на станции Дмитров (LE), в 1973 году — в Рязанской области (город Касимов по склону оврага, в бурьяне. 1973. В. Тихомиров, Н. Октябрева. MW), где уже через 2 года отмечена как компонент флоры Рязанской Мещеры [Тихомиров, 1975]. В 1976 году недотрога найдена в Брянской области (МНА). При изучении флоры Калининской области I. glandulifera найдена на пустырях, особенно во влажных местах (в канавах, около прудов, в понижениях рельефа) в Калинине, Торжке, Старице, Кувшинове, Нелидове, Западной Двине, Торопце [Малышева, 1979]. В 1980 году заросли цветущих растений найдены в городе Чаплыгин Липецкой области в пойме реки Становая Ряса, годом позднее в городе Липецк также в большом количестве, а в 1987 году — в поймах реки Воронеж и Дон [Вьюкова, 1983, 1987]. К 1982 году относятся сборы в Орловской области (город Орёл, стройка у сараев. 11.08.1982 год В. И. Радыгина. ОГУ). В 1983 году недотрога встречена во многих районах и пригородах Рязани [Гущина, 1983], а в 1988 году — в Ульяновской области (МНА). В 1989 году I. glandulifera обнаружена во влажных местообитаниях на берегу Воронежского водохранилища [Барабаш, Камаева, 1989] и занесена в Марийскую АССР [Абрамов, Киряков, 1989]. В настоящее время широко распространена во всех областях Средней России.
Местообитания. В природе I. glandulifera растёт на высоте 1 800-3 000 метров, часто по берегам рек, по краям полей и в нарушенных влажных лесах [Valentine, 1971], а в Пакистане — до 4 300 метров над уровнем моря (Nasir, 1980).
В пределах вторичного ареала произрастает в широком диапазоне местообитаний, но лучше всего растёт на сырых местах, богатых питательными веществами, особенно по берегам озёр, по рекам и ручьям. Часто вид также можно обнаружить в антропогенно нарушенных и искусственных местообитаниях — на полях, в кустарниках, канавах, обочинах дорог и живых изгородях [Kurtto, 1996]. Наиболее подходящими для I. glandulifera являются биотопы, подверженные антропогенному воздействию [Garkaje, 2006]. В таких биотопах I. glandulifera обычно растёт вместе с растениями, которые нуждаются в запасе азота в почве — Aegopodium podagraria, Urtica dioica, Calystegia septum, Deschampsia caespitosa, Stellaria nemorum и Galium aparine. Вид вторгается в многолетнюю травянистую растительность речных берегов, светлые пойменные леса и влажные луга и найден на разнообразных типах почвы [Kowarik, 2003].
В Европе растения I. glandulifera всех возрастов нехолодостойки. Обычно все взрослые растения погибают осенью при первых морозах, а проростки погибают при поздних весенних заморозках [Sebald et al., 1998]. Вид также не выносит засухи и быстро увядает, и растения могут выживать, только если период засухи короток [Beerling, Perrins, 1993].
Недотрога желёзконосная — теневыносливое растение. Рост её, однако, ухудшается, когда мощность светового потока становится менее 30% полного дневного света [Beerling, Perrins, 1993]. В качестве факторов, которые ограничивают инвазию I. glandulifera, могут выступать требовательность к высокой влажности почвы и слабая морозоустойчивость. Этот вид отмечен как на равнинных участках, так и на крутых (>40°) склонах, при условии умеренного затенения, способен произрастать как на тонком, так и на грубом аллювии, морской гальке, хорошо дренируемых минеральных почвах, отвалах каменного угля и торфах [Beerling, Perrins, 1993]. Недотрога желёзконосная обычно растёт на богатых минеральным азотом почвах, предпочитает почвы с нейтральной реакцией, но может расти на кислых и щелочных (рН от 4,5 до 7,3-7,7). Обитает на почвах от влажных, не просыхающих, до сырых, часто плохо аэрируемых. Для успешного выживания проростков требуются умеренная нарушенность растительного покрова (вывалы деревьев, упавшие ветки деревьев, зимнее подтопление берегов) и наличие голых участков грунта [Марков и другие, 1997].
В Московской области I. glandulifera встречается на берегу реки Москва на протяжении десятков километров в Можайском, Рузском и Одинцовском районах. Местами заходит на ключевые болота. В ряде районов вид образует обширные заросли по заболоченным лугам у водоёмов [Игнатов и другие, 1990]. Следовательно, во вторичном ареале I. glandulifera занимает те же местообитания, что и на родине.
Габитус растений меняется в зависимости от влажности местообитания. Высокие, маловетвистые экземпляры обитают в довольно влажных условиях, низкие растения с обильно ветвящимися стеблями, короткими междоузлиями и мелкими листьями характерны для нарушенных лесных участков с непостоянным увлажнением [Марков и другие, 1997]. Плотность популяций в среднем составляет 40 растений/м2, но может возрастать и до 70 особей [Wadsworth et al., 2000]. Достижение высокого роста растений при низком уровне освещения в плотных популяциях обеспечивается физиологическим процессом накопления в клетках стебля аниона NO3 [Andrews et al., 2005].
Размножение и жизненный цикл. Цветки I. glandulifera протандричные, поэтому опыление их облигатно перекрёстное. Однако может иметь место опыление между цветками с одного и того же растения. Вид энтомо-фильный, опыляется шмелями.
I. glandulifera — однолетнее растение. В зависимости от плотности популяции и от характеристики места произрастания одна особь может продуцировать от 95 до 390 плодов и от 500 до 2 500 семян [Марков и другие, 1997]. Обычно семена выбрасываются из коробочки на расстояние до 5 м, максимальное число семян падает на почву в радиусе 1-2 м от материнского растения. Подсчитано, что скорость распространения популяции в одном местообитании составляет 2,47 м в год [Beerling, Perrins, 1993]. В переувлажнённых местах набухшие семена I. glandidifera погружаются в воду и способны прорастать под водой [Lhotska, Kopecky, 1966]. Более высокие растения образуют большее число плодов и большее число семян в одном плоде. Масса семян не зависит от числа плодов на растении или от числа семян в плоде. Масса 100 семян варьирует незначительно [Виноградова, 2008] и составляет в среднем от 1,05 г (Санкт-Петербург) до 1,59 г (Вологда). На высокопитательных почвах I. glandulifera продуцирует большее количество семян, а не более крупные семена [Willis, Hulme, 2004].
В Англии семена могут прорастать иногда через 18 месяцев. Без холодной стратификации семян большинство сеянцев появляется в течение 4 недель [Beerling, Perrins, 1993]. По нашим данным, семена I. glandulifera теряют всхожесть уже через 1 год сухого хранения.
I. glandulifera имеет хорошую регенерационную способность, и на повреждённых стеблях формируются новые ветви и цветки. Мелкие растения также могут образовывать цветки и семена.
В Германии время от прорастания до начала цветения — 13 недель, а цветение продолжается в течение следующих 12 недель [Sebald et al., 1998].
Расселение и распространение. I. glandulifera распространяется только семенами. Распространение семян происходит гидро-, антропо- и автохорно. Когда коробочка созревает, она взрывается и выбрасывает семена. Имеются сообщения, что семена рассеиваются от материнского растения на расстояние до 7 м. Отдельное растение может производить более 4 тысяч семян, и в чистых зарослях образуется до 32 тысяч семян/м2 [Koenies, Glavac, 1979]. Экспансия вида в речных системах, в частности, обусловлена способностью семян к распространению по водным путям, поскольку они могут переноситься и плавно текущей водой (в осадке), и сухими плавучими семенами. Вместе с наносами сезонных паводков семена могут переноситься вдоль рек на большие расстояния. Мелкие семена легко переносятся в новые местообитания с почвой или на обуви. Семена, вероятно, также распространяются муравьями (мирмекохория). Плодоносящие экземпляры или их фрагменты также переносятся с почвой или потоками воды [Kurtto, 1993]. Для Великобритании рассчитана способность к расселению, которая составляет 2,6-5 км в год (ссылка).
Репродуктивная стратегия основана на активном распространении семян и на их высокой всхожести, которая составляет до 80% [Grime, 1987]. Растения конкурируют на речных берегах путём синхронного прорастания множества семян, быстро вырабатывают достаточную биомассу и подавляют соседние виды. I. glandulifera растёт довольно быстро и формирует плотные заросли.

Внедрение Impatiens glandulifera в прибрежные сообщества, Московская область, река Москва в Одинцовском районе
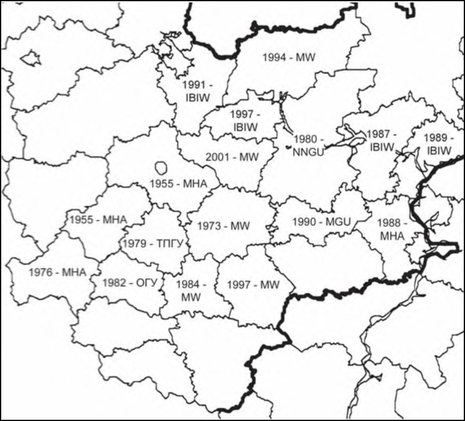
Расселение Impatiens glandulifera в Средней России

Массовое цветение Impatiens glandulifera в Москве, в овраге в Битцевском лесопарке

Розовая окраска венчика Impatiens glandulifera

Дурнишник эльбский
Xanthium albinum (Widder) H.Scholz

Недотрога мелкоцветковая
Impatiens parviflora DC.




