Вторичный ареал: Золотая розга гигантская, или Золотарник гигантский
Solidago gigantea Ait.
Вторичный ареал
История интродукции и географическое распространение. Впервые как декоративное интродуцированное растение S. gigantea зарегистрирована в Ботаническом саду Лондона в 1758 году [Weber, 1998]. Вскоре она была отмечена в садах и питомниках континентальной Европы, но происхождение этих инициальных популяций остаётся неизвестным. В первой половине XIX века S. gigantea зарегистрирована во Франции, в Швейцарии и Германии [Weber, Jakobs, 2005]. Вид стал инвазионным через ~100 лет: первое упоминание о самопроизвольном распространении в Германии датируется 1832 годом, в Австрии — 1857 год [Wagenitz, 1979]. Между 1850 и 1880 годами описано много новых местонахождений, и растение начало распространяться по всей Центральной Европе [Wagenitz, 1979]. На основании гербарных образцов и литературных источников, можно утверждать, что скорость экспансии составляла ~910 км2/год [Weber, 1998]. В 1950 году S. gigantea уже распространилась по всей Европе от севера Испании до Восточной Европы и от Северной Италии до юга Скандинавии [Tutin et al., 1964]. В настоящее время ареал вида в Европе простирается от 42° до 63°северной широты и ожидается дальнейшее расширение его границ в восточном направлении [Meusel, Jager, 1992; Weber, 2001]. S. gigantea встречается, главным образом, ниже 1 200 метров надо уровнем моря, но может иногда наблюдаться на больших высотах [Weber, Jakobs, 2005].
В европейской части ареала отмечено локальное разнообразие фенотипов: внутривидовые формы широко распространены в Центральной и Восточной Европе, но значительно реже встречаются в Скандинавии и Северной Германии. По мнению ряда учёных [Botta-Dukat, Dancza, 2001а], именно высокий уровень изменчивости морфологических признаков обеспечил столь высокий инвазионный потенциал вида в Европе.
Помимо Европы, S. gigantea также натурализовалась в Японии, Корее [Enomoto, 2000; Tsuyuzaki, 2002] и на Дальнем Востоке РФ [МНА; Баркалов, 1992], в Новой Зеландии, Австралии, на Азорских и на Гавайских островах, а также в Мексике [Weber, Jakobs, 2005].
Пути и способы заноса. S. gigantea интродуцирована в Европу как декоративное растение и пользовалась большим успехом у садовников из-за лёгкости выращивания. Культивирование золотой розги на приусадебных участках до сих пор является основным путём заноса вида на новые территории.
При натурализации S. gigantea вначале захватывает рудеральные нарушенные территории — берега рек и обочины автомобильных дорог и ж/д полотна. С конца XIX века S. gigantea также закрепилась в полуестественных и естественных местообитаниях [Guzikowa, Maycock, 1986]. Урбанизация, строительство автомобильных дорог и ж/д путей, заброшенные поля способствуют распространению вида. Избавление от садового мусора (удаление растений с дачных участков) и выращивание золотой розги как медоносного растения также содействуют его инвазии в новые места обитания [Weber, Jakobs, 2005]. По сравнению с другими видами золотарника, интродуцироваными в Европе, скорость распространения S. gigantea оказалась наиболее высокой [Guzikowa, Maycock, 1986; Weber, 1998], и сегодня вид обоснованно относится к одним из наиболее агрессивных инвазионных растений Европы.
В 1937 году на участках Интродукционного питомника субтропических культур в городе Сухуми, куда Н. И. Вавиловым были завезены в Россию вместе с перспективным каучуконосом S. leavenworthii Torr. et Gray семена других золотарников, было представлено уже 50 видов Solidago L., в том числе 1 образец S. gigantea [Волховская,1937].
Статус в регионе. Как интродуцированное растение отмечено в Воронежской области в 1868 году [MW]. В настоящее время как декоративное и изредка дичающее растение встречается по всей Средней России. В естественные ценозы активно внедряется в последнее десятилетие. Массовые заросли отмечены в Калужской, Курской и Московской областях.
Местообитания. В естественном ареале S. gigantea является частью сообществ прерий и заливных лугов. Это высоко-толерантный вид, не требовательный к свету, влажности почвы, содержанию питательных веществ, температуре и рН-фактору [Weber, Jakobs, 2005]. Хотя S. gigantea предпочитает богатые и достаточно увлажнённые почвы, она встречается на различных по структуре грунтах и на разных типах почв. Наилучшего развития S. gigantea достигает в прибрежных местообитаниях и на топких местах, но растёт также и в сухих местах, таких как обочины автомобильных дорог и ж/д насыпи.
Во вторичном ареале в Европе S. gigantea занимает схожие экологические ниши. Вид произрастает, главным образом, в местообитаниях, богатых азотом, но также найден на относительно бедных питательным веществом участках — пустырях и обочинах дорог. На увеличение питательных веществ в почве вид отвечает значительным увеличением размеров, особенно размера соцветий и числа произведённых семян. В сырых и влажных местообитаниях её жизненность выше, чем в сухих местах, и часто этот вид образует плотные одновидовые насаждения. Однако продолжительного затопления этот вид не выносит. В более сухих местах S. gigantea часто сосуществует с другими видами [Botta-Dukat, Dancza, 2001а], и в таких условиях вид менее конкурентоспособен. S. gigantea реагирует на фактор сухости уменьшением листовой поверхности и редукцией общей биомассы. Механическое удаление корневищ может сократить доминирование S. gigantea в таких местообитаниях, поскольку скорость побего-образования замедляется [Botta-Dukat, Dancza, 2001b],
Размножение и жизненный цикл. S. gigantea — геофит, внутри клонов практически не наблюдается сеянцев, а потому исследователи предполагают, что семена служат, главным образом, для распространения на дальние расстояния и заселения новых мест. У близкородственного вида S. altissima внутри клонов прорастание семян ингибируется дитерпеноидами, выделяемыми растениями, подобное может иметь место и в случае S. gigantea [Werner et al., 1980]. Иногда при наличии открытых участков сеянцы могут появляться после весеннего покоса [Weber, Jakobs, 2005]. Однако размер популяции обычно увеличивается за счёт клонального роста. Клон растёт, главным образом, в течение первых лет после возникновения, а по мере его старения роль полового размножения возрастает.
Семеношение наблюдается каждый год, хотя невысокие побеги внутри клона могут оставаться вегетативными. С другой стороны, у изолированно растущих растений в неблагоприятных условиях могут цвести даже небольшие побеги высотой до 25 см. В благоприятных условиях почти все растения цветут и плодоносят на втором году жизни. S. gigantea цветёт поздно, с июля по ноябрь, с пиком цветения в августе — сентябре. Вид является перекрёстно-опылителем: соцветия привлекают опылителей яркими жёлтыми цветами и большим количеством пыльцы и нектара. Частые посетители цветов в Европе — виды таких семейств, как Phalacridae, Muscidae, Syrphidae, Apidae, Formicidae, Sphecidae и Panorpidae [Weber, Jakobs, 2005].
Отрастание побегов начинается весной, и высота их увеличивается почти линейно до конца июля. Растения заканчивают рост только после формирования соцветия и раскрывания первых цветков, обычно между августом и сентябрём. В благоприятных условиях отдельный генеративный побег продуцирует до 19 тысяч семянок. Семена легко разносит ветер, но большинство семянок остаются на стебле в течение зимы. Семянки прорастают весной, но период прорастания довольно растянут и продолжается до начала лета. Всхожесть семян S. gigantea может составлять 100%, однако этот параметр зависит от почвенных условий: на глинистой почве и на торфе всхожесть составила 47%, тогда как на фильтровальной бумаге в течение 3 недель проросло 99% семян [Weber, Jakobs, 2005]. В условиях ограниченных ресурсов половое размножение сокращается, а преобладать начинает вегетативное размножение.
Расселение и распространение. S. gigantea может увеличивать захваченные площади как с помощью семян, которые ветер разносит на далёкие расстояния, так с помощью фрагментов корневищ, которые переносятся водными потоками и легко укореняются. Распространение вдоль водного пути часто может быть заметным; например, популяция золотой розги легко распространилась вдоль реки Тальяменто на севере Италии [Weber, Jakobs, 2005].
Имеется отрицательная зависимость между массой семян и их численностью, и соотношение этих величин зависит от конкретных почвенно-климатических условий. В менее благоприятных условиях и вытаптываемых (часто нарушаемых) местах семян формируется больше, и они легче, что гарантирует их перенос на дальние расстояния; там, где условия благоприятнее, семян меньше, но они крупнее, а это повышает вероятность образования сеянцев в близлежащих местах с сомкнутым растительным покровом [Werner, Piatt, 1976].
Корневища растут в направлении от центра клона. Корневища довольно длинные и сильно ветвятся, таким образом, популяции состоят из нескольких или многих смешивающихся генет. Соотношение генеративных и вегетативных побегов существенно меняется в зависимости от почвы и условий освещённости. Отмечено [Weber, Jakobs, 2005], что число генеративных побегов отрицательно коррелирует с плотностью побегов в популяции. В центре зарослей сопутствующие виды практически отсутствуют, по направлению к краям число сопутствующих видов повышается [Lukac, 1988]. На некосимом заливном лугу в Швейцарии площадь популяции за счёт роста клонов увеличивалась на 0,3-0,8 м/год [Weber, Jakobs, 2005]. В одной рамете за вегетационный сезон формируется от 5 до 55 корневищ. Большое число почек на корневищах позволяет растениям быстро повторно отрастать после повреждений. Число ростовых почек на корневищах у S. gigantea в 10 раз выше, чем у S. canadensis [Weber, Jakobs, 2005], и это в какой-то мере объясняет успешность колонизации S. gigantea. Кроме того, корневища S. gigantea в среднем намного более длинные; таким образом, вид может покрыть большие площади и расширить размер участка, захваченный популяцией, за относительно более короткий промежуток времени [Schmid et al., 1988]. При отсутствии травоядных животных корневища остаются в виде неделимой системы в течение нескольких лет.
Вероятно, отсутствие травоядных животных во вторичном ареале может быть одной из причин более высокого числа корневищ, отмечаемых в инвазионных популяциях. Размер площади заросли S. gigantea зависит от количества осенних осадков [Weber, Jakobs, 2005]. В норме длина корневищ составляет до 90 см. В благоприятных условиях корневища в среднем короче, и тогда клоны более плотные; более длинные корневища, которые развиваются в менее благоприятных условиях, имеют больше шансов достигнуть подходящих участков. На недавно заброшенных полях индивидуальные клоны часто легко различимы и достигают диаметра 2-5 м. Крупные старые клоны могут состоять из 1 тысячи рамет. Иногда клоны формируют «ведьмины круги» вокруг начального центра генеты.
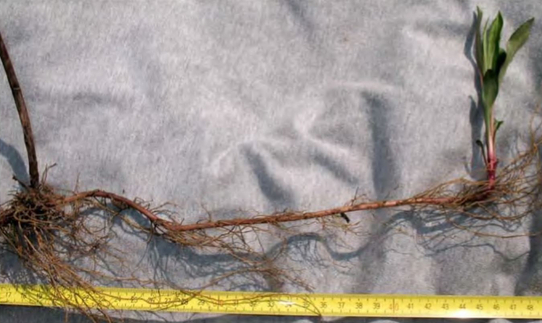
Корневища Solidago сanadensis (вверху) и Solidago gigantea (внизу)

Соцветия Solidago gigantea (слева) и Solidago canadensis (справа)
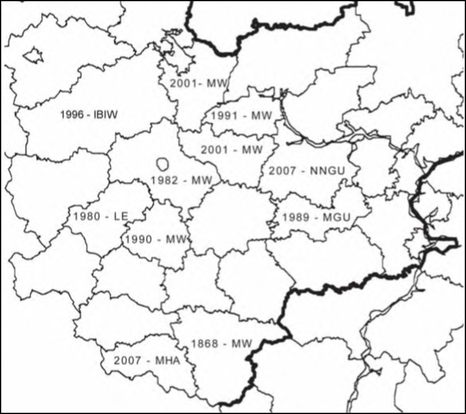
Расселение Solidago gigantea в Средней России

Внедрение Solidago gigantea в прибрежные сообщества

Золотая розга канадская, или Золотарник канадский
Solidago сanadensis L.

Дурнишник эльбский
Xanthium albinum (Widder) H.Scholz




